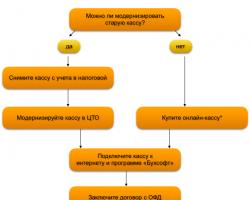
Билет 15
1. Формы (фрагменты) афферентного синтеза: Доминирующая мотивация; Обстановочная афферентация; Пусковая афферентация. Роль ретикулярной формации.
2. Быстрые и медленные мышечные волокна.
Вопрос 1
АФФЕРЕНТНЫЙ СИНТЕЗ - (соединение, составление) - процесс сопоставления, отбора и синтеза многочисленных и разных по функциональному значению афферентаций, вызванных различными воздействиями на организм, происходящий в ц. н. с., на основе к-рого формируется цель действия.
А. с. согласно теории функциональной системы Анохина - первая, универсальная, стадия любого целенаправленного поведенческого акта (см. Функциональные системы).
А. с. включает обработку 4 основных видов афферентных возбуждений.
1. Мотивационное возбуждение отражает доминирующую потребность организма, к-рая возникает под влиянием метаболических, гормональных, а у человека - и социальных факторов Мотивация играет решающую роль в формировании цели действия. Специфически повышая реактивность корковых нейронов с помощью ориентировочно-исследовательской реакции , мотивационное возбуждение способствует обработке и активному отбору сенсорной информации, необходимой для построения целенаправленного поведения.
2. Обстановочная афферентация представляет собой воздействие на организм всей совокупности внешних факторов, составляющих конкретную обстановку, на фоне к-рой развертывается приспособительная деятельность. Обстановочная афферентация формируется не только постоянными компонентами обстановки, но и рядом последовательных афферентных воздействий на организм. Характерная особенность обстановочной афферентации состоит в том, что она придает специфику будущей поведенческой реакции, обеспечивая ее приспособительное значение только в данной обстановке.
Наиболее отчетливо роль обстановочной афферентации проявляется в опытах с условными рефлексами. В этих случаях на один и тот же условный раздражитель животное отвечает условной оборонительной реакцией в одной экспериментальной камере и условной пищевой - в другой (или в одной и той же экспериментальной камере утром животное отвечает пищевой реакцией, а вечером - оборонительной).
На стадии афферентного синтеза решаются вопросы “что делать?”, “как делать?”, “когда делать?”.
Пусковая афферентация
Представляет собой специальный раздражитель, собственно запускающий поведенческую реакцию. Значение пускового раздражителя состоит в том, что он призван обозначить момент начала поведенческой реакции.
Целенаправленное поведение может начинаться и без явного пускового стимула. Примерами таких реакций являются регулярно совершающиеся физиологические отправления (еда, сон, дефекация, мочеиспускание и др.), приуроченные к определенным периодам суток.
Афферентный синтез осуществляется на основе следующих нейрофизиологических механизмов:
1) механизмы восходящих активирующих влияний подкорковых образований на кору головного мозга. Это прежде всего, активирующие влияния гипоталамуса к лобным отделам коры, через передние ядра таламуса, что отражает мотивационные возбуждения. Аналогичным образом воздействуют другие лимбические системы. Вторыми по активирующему значению являются ретикулярные структуры среднего мозга и моста, которые обеспечивают соответствующий уровень бодрствования.
2) механизмы конвергенции возбуждений различного качества на нейронах коры и подкорковых структурах мозга. В частности, мультисенсорная конвергенция от поверхностей (зрительная, тактильная, слуховая, температурная и др.); мультибиологическая конвергенция, связанная с определенными состояниями (голод, боль и т.п.) и др.;
3) интеграцию мотивационных, обстановочных и пусковых афферентаций на нейронах коры мозга;
4) механизмы формирования доминанты, за счет которых подавляется текущая деятельность и удерживается вновь сформированная поведенческая реакция.
Роль ретикулярной формации
Ретикулярная формация характеризуется относительно малой возбудимостью. Эффекты ее раздражения появляются через большой латентный период, она медленно реагирует и остается активной в течение продолжительного времени после прекращения раздражения (длительное последействие). Ретикулярная формация облегчает или подавляет фазные движения и напряжение скелетных мышц, вызываемые мотонейронами спинного мозга, а также движения, вызванные с коры больших полушарий. Ретикулярная формация среднего и промежуточного мозга облегчает рефлекторные движения животных, раздражение промежуточного мозга тормозит двигательные рефлексы спинного мозга.
Боковые отделы ретикулярной формации варолиева моста и среднего мозга облегчают, а средние ее отделы в продолговатом мозге тормозят двигательные рефлексы. Облегчение и торможение зависят также от интенсивности и продолжительности раздражения ретикулярной формации. По гамма-нейронам она регулирует функции мышечных веретен, следовательно, обратную информацию из скелетных мышц. Она изменяет также возбудимость восходящих афферентных путей спинного мозга, что может снизить или прекратить постсинаптическое торможение. Тонические влияния ретикулярной формации вызывают ВПСП или ТПСП в мотонейронах спинного мозга. Она изменяет также передачу импульсов в мозговом стволе и одновременно с влиянием на скелетную мускулатуру вызывает сосудодвигательные, дыхательные, зрачковые и другие реакции.
Ретикулярная формация оказывает адаптационно-трофическое влияние на кору больших полушарий, подкорковые образования промежуточного мозга, мозжечок и спинной мозг. Существуют взаимные влияния этих отделов нервной системы, как возбуждающие, так и тормозящие. Она участвует в физиологических процессах сна и пробуждения, а также в эмоциях, в реакции напряжения («стресс») и др. Раздражение ретикулярной формации вызывает пробуждение спящих животных, а ее разрушение и выключение - глубокий сон у бодрствующих животных. Изучены взаимные влияния ретикулярной формации и коры больших полушарий.Установлено участие ретикулярной формации в образовании и протекании условных рефлексов
По симпатическим волокнам ретикулярная формация регулирует возбудимость и работоспособность скелетной мускулатуры, функциональное состояние нервной системы и органов чувств, оказывая на них адаптационно-трофическое влияние. Регуляция рефлексов позы и двигательных рефлексов, перемещающих тело, осуществляется по эфферентным гамма-волокнам иннервирующим проприоцепторы.
Ретикулярная формация регулирует вегетативные функции, деятельность внутренних органов. Она влияет на образование гормонов в гипофизе и других железах внутренней секреции и в ней концентрируются гормоны и медиаторы.
Афферентные волокна поступают в нее по симпатическим и блуждающим нервам. Частьклеток ретикулярной формации среднего мозга и варолнева моста возбуждается адреналином и норадреналином (адренореактивные системы) а другая часть, расположенная в промежуточном мозге, несколько выше среднего мозга, возбуждается ацетилхолином и его производными (холинореактивные системы). Адренореактивные системы среднего мозга и варолиева моста облегчают наступление двигательных рефлексов, а адренореактивные системы продолговатого мозга тормозят спинномозговые рефлексы. Адреналин возбуждает и холинореактивные системы. Предполагается, что действие ацетилхолина и его производных менее ограничено, чем действие адреналина, и охватывает многие области головного мозга. Действие ацетилхолина на ретикулярную формацию противоположно его периферическому влиянию на внутренние органы. Ретикулярную формацию среднего и продолговатого мозга возбуждает углекислота.
Гормоны и медиаторы действуют на функцию больших полушарий как непосредственно, так и посредством ретикулярной формации. Таким образом, ретикулярная формация мозгового ствола - подкорковый центр вегетативной нервной системы.
Вопрос 2 .
Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:

Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
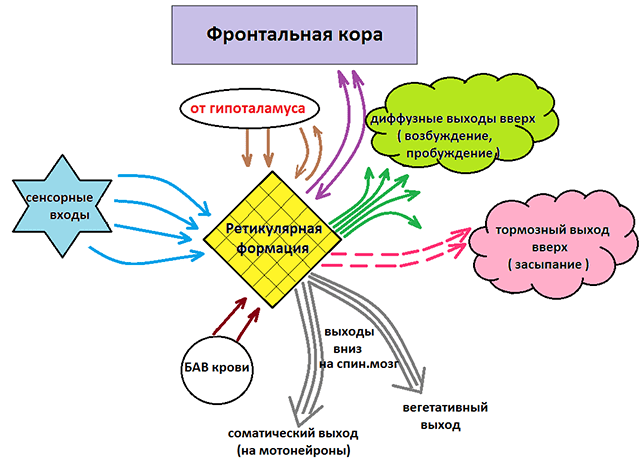
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
Ретикулярная формация известна с 1845 г., описана Дейтерсом (O.F.C. Deiters) в 1885 г. В настоящее время продолжается ее изучение. Ретикулярная формация располагается между задними и боковыми рогами шейных сегментов спинного мозга, в покрышке мозгового ствола, в центральном ядре зрительного бугра. Представляет собой комплекс анатомически и функционально взаимосвязанных нейронов, окруженных множеством волокон, идущих в различных направлениях к ядерным структурам и проводящим путям (рис. 30).
Рис. 30. : 1 - восходящие пути; 2 - нисходящие пути; 3 - специфический (лемнисковый) чувствительный проводящий путь; 4 - пирамидальный путь.
Ретикулярная формация воспринимает все импульсы (болевые, температурные, световые, звуковые и пр.), однако в ней нет специализированных нейронов. Поэтому одни и те же нейроны воспринимают различные импульсы и передают их в различные отделы мозга, во все участки коры. Ретикулярная формация - это вторая афферентная система головного мозга, его неспецифическая структура . Она имеет двухсторонние связи со всеми структурами головного и спинного мозга (рис. 31, 32).
 Рис. 31. : 1; 2; 3 - специфический (лемнисковый) чувствительный проводящий путь; 4 - коллатерали, соединяющие специфический чувствительный путь с ретикулярной формацией мозгового ствола; 5 - восходящая активирующая система ретикулярной формации; 6 - генерализованное влияние ретикулярной формации на кору головного мозга.
Рис. 31. : 1; 2; 3 - специфический (лемнисковый) чувствительный проводящий путь; 4 - коллатерали, соединяющие специфический чувствительный путь с ретикулярной формацией мозгового ствола; 5 - восходящая активирующая система ретикулярной формации; 6 - генерализованное влияние ретикулярной формации на кору головного мозга.
 Рис. 32. : 1 - чувствительный нерв, на который наносится стимул (болевое раздражение); 2 - спинной мозг; 3 - симпатические нервы; 4 - надпочечник; 5 - каротидный синус; 6 - гипофиз; 7 - ретикулярная формация. Сплошными стрелками обозначены нервные влияния, пунктирными - гормональные влияния, которые через ретикулярную формацию оказывают активирующее воздействие на кору головного мозга.
Рис. 32. : 1 - чувствительный нерв, на который наносится стимул (болевое раздражение); 2 - спинной мозг; 3 - симпатические нервы; 4 - надпочечник; 5 - каротидный синус; 6 - гипофиз; 7 - ретикулярная формация. Сплошными стрелками обозначены нервные влияния, пунктирными - гормональные влияния, которые через ретикулярную формацию оказывают активирующее воздействие на кору головного мозга.
Структурные элементы ретикулярной формации ствола мозга подразделяют на латеральный и медиальный отделы. В латеральном отделе заканчиваются волокна из различных афферентных систем. К рассеянным клеткам и ядрам ретикулярной формации подходят коллатерали от медиальной и латеральной петель, от чувствительных ядер черепных нервов. От нейронов медиального отдела начинаются эфферентные волокна к двигательным ядрам черепных нервов, к мозжечку, к двигательным ядрам передних рогов спинного мозга.
Основные афферентные пути ретикулярной формации: tr. spinoreticularis - от спинного мозга, tr. tegmentothalamicus - от среднего мозга, reticulothalamicus - от продолговатого мозга и моста, tr. thalamocorticalis - ко всем областям и слоям коры полушарий большого мозга. Сетевидная формация активизирует кору большого мозга и мозжечок.
Кора полушарий большого мозга, в свою очередь, посылает по tr. corticoreticularis импульсы в ретикулярную формацию в составе пирамидных путей. Основным эфферентным трактом является tr. reticulospinalis. Этот путь проводит тонические импульсы к гамма-мотонейронам спинного мозга. Ретикулярная формация регулирует моторное звено, обеспечивая координацию движений, синхронность мышечных сокращений, обеспечивает нестандартные движения, рефлекс равновесия, устанавливает антигравитационный тонус мускулатуры, удерживающий тело над землей. Ретикулярная формация перераспределяет мышечный тонус, что в кризисных ситуациях приводит к мобилизации скрытых резервов организма.
Установлена роль голубоватого пятна и ядер шва в регуляции сна и бодрствования. Голубоватое пятно (locus caeruleus ) находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют норадреналин, активирующий вышележащие отделы мозга. Особенно высока активность нейронов голубоватого пятна во время бодрствования, во время глубокого сна она угасает почти полностью.
Ядра шва (nuclei raphes ) располагаются по срединной линии продолговатого мозга. Нейроциты этих ядер вырабатывают серотонин, который вызывает процессы разлитого торможения и состояние сна.
Ядра ретикулярной формации продолговатого мозга имеют связи с вегетативными ядрами Ⅸ, Ⅹ нервов и симпатическими ядрами спинного мозга. Поэтому они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и так далее.
Ядра Кахаля и Даркшевича, относящиеся к ретикулярной формации среднего мозга, к медиальному продольному пучку (fasciculus longitudinalis medialis ), имеют связи с ядрами третьей, четвертой, шестой, восьмой, девятой, десятой и одиннадцатой парой черепных нервов. Они координируют работу этого пучка, обеспечивая сочетанные повороты головы и глаз при изменении позы или при поиске источника звука, фиксацию взгляда. (Данные движения совершенно необходимы при трудовых и игровых актах).
Эти связи объясняют вегетативные нарушения при вестибулярных перегрузках. Рассеянные нейроны ретикулярной формации выступают в качестве вставочных нейронов охранительных рефлексов глотательного, роговичного (рис. 33), кашлевого рвоты, зевоты, чихания и др.
 Рис. 33. : 1 - рецепторы, расположенные в роговице; 2 - глазная ветвь тройничного нерва; 3 - ложноуниполярная клетка чувствительного узла тройничного нерва; 4 - ассоциативный нейрон - рассеянная клетка ретикулярной формации; 5 - клетка двигательного ядра лицевого нерва; 6 - круговая мышца глаза.
Рис. 33. : 1 - рецепторы, расположенные в роговице; 2 - глазная ветвь тройничного нерва; 3 - ложноуниполярная клетка чувствительного узла тройничного нерва; 4 - ассоциативный нейрон - рассеянная клетка ретикулярной формации; 5 - клетка двигательного ядра лицевого нерва; 6 - круговая мышца глаза.
Филогенетически очень древняя нейронная структура и хорошо развитый отдел ствола мозга рептилий. Сначала она представляла собой полисинаптический путь с медленным проведением, тесно связанный с обонятельной и лимбической областями. Прогрессирующее доминирование зрения и слуха над обонянием привело к смещению чувствительных и двигательных функций внутрь покрышки среднего мозга. Прямые спино-покрышечный и покрышечно-спинномозговой пути обходят ретикулярную формацию, которая отвечает, главным образом, за вегетативную регуляцию. У млекопитающих покрышка, в свою очередь, стала играть второстепенную роль в передаче возбуждения по очень быстро проводящим волокнам, соединяющим кору полушарий с периферическими двигательными и чувствительными нейронами.
В мозге человека ретикулярная формация сохраняет свою связь с лимбической системой и продолжает играть важную роль в вегетативной и рефлекторной регуляции.
Термин ретикулярная формация относят только к полисинаптической нейронной сети ствола мозга, несмотря на то, что сеть распространяется кпереди в таламус и гипоталамус и кзади в проприоспинальный тракт спинного мозга.
Общее строение показано на рисунке ниже. Срединная ретикулярная формация образована рядом ядер шва (греч.-nuclei raphe). Большая часть серотонинергических путей осевого отдела нервной системы начинается из ядер шва.
Ретикулярная формация (РФ).Рядом расположена парамедианная ретикулярная формация. Этот отдел полностью состоит из крупноклеточных нейронов; в нижнем отделе моста и верхнем отделе продолговатого мозга (до уровня сращения ретикулярной формации с центральным ретикулярным ядром продолговатого мозга) можно также обнаружить гигантоклеточные нейроны.
Самым передним отделом считают латеральную мелкоклеточную ретикулярную формацию . Длинные дендриты мелкоклеточных нейронов образуют разветвления через определенные интервалы. Дендриты имеют преимущественно поперечное направление, а через промежутки между ними проходят длинные проводящие пути к таламусу. Латеральный отдел образован, главным образом, афферентными нейронами. К ним подходят волокна от всех чувствительных проводящих путей, включая органы чувств.
Обонятельные волокна проходят через медиальный пучок переднего мозга, расположенный рядом с гипоталамусом.
Зрительные проводящие пути проходят через верхний холмик.
Слуховые волокна подходят от верхнего ядра оливы.
Вестибулярные волокна подходят от медиального вестибулярного ядра.
Соматические чувствительные волокна проходят через спинно-ретикулярные тракты от спинномозгового и собственного (главного или главного мостового) ядер тройничного нерва.
Большая часть аксонов мелкоклеточных нейронов интенсивно разветвляется между дендритами нейронов парамедианной ретикулярной формации. Однако часть из них образует синапсы с ядрами черепных нервов и участвует в создании программ движений.
Парамедианная ретикулярная формация - преимущественно эфферентная система. Аксоны относительно длинные, некоторые поднимаются вверх, образуя синапсы с ретикулярной формацией ствола мозга или таламусом. От других отходят как восходящие, так и нисходящие ветви, образующие полисинаптическую сеть. К крупноклеточным нейронам подходят волокна от премоторной коры, которые дают начало ретикуло-спинномозговым путям моста и продолговатого мозга.

а) Аминергические нейроны ствола мозга . Рассеянные по ретикулярной формации группы аминергических (или моноаминергических) нейронов - нейроны, медиаторы которых образуются из ароматических аминокислот и обладают рядом влияний на клетку. Одна группа продуцирует нейромедиатор серотононин, три другие - катехоламины (дофамин, норадреналин и адреналин), одна группа - гистамин.
 Серотонинергические пути от срединного отдела ствола мозга (шва).
Серотонинергические пути от срединного отдела ствола мозга (шва).
Серотонинергические нейроны - наиболее часто встречаемые нейроны в любом отделе центральной нервной системы (ЦНС). К ним относят нейроны среднего мозга, волокна которых поднимаются к большим полушариям; нейроны моста, разветвляющиеся в стволе мозга и мозжечке; клетки продолговатого мозга, нисходящие в спинной мозг.
Все отделы серого вещества ЦНС пронизаны серотонин-секретирующими аксональными ветвями. Повышение серотонинергической активности используют в клинической практике для лечения такого распространенного заболевания, как большое депрессивное расстройство.

Дофаминергические нейроны среднего мозга представлены двумя группами. В месте соединения покрышки с ножками расположено черное вещество. Медиальнее него расположены вентральные ядра покрышки, от которых отходят мезокортикальные волокна к лобной доле и мезолимбические волокна, идущие непосредственно к прилежащему ядру.
Норадренергические (норэпинефринергические) нейроны немного менее многочисленные, чем серотонинергические. Около 90 % тел нейронов сконцентрировано в голубом пятне (locus ceruleus) в дне IV желудочка у верхнего конца моста. От голубого пятна начинаются пути во всех направлениях, что показано на рисунке ниже.
 Норадренергические пути от моста и продолгова того мозга.
Норадренергические пути от моста и продолгова того мозга.
Адреналин-секретирующие (эпинефрин-секретирующие) нейроны относительно немногочисленны и расположены преимущественно в ростральном/каудальном отделах продолговатого мозга. Одна часть волокон восходит к гипоталамусу, другая направляется книзу, образуя синапсы с преганглионарными симпатическими нейронами спинного мозга.
В больших полушариях ионная и электрическая активность аминергических нейронов значительно различается. Во-первых, для каждого амина существует более одного типа постсинаптических рецепторов. Во-вторых, некоторые аминергические нейроны высвобождают также белковые вещества, способные регулировать действие медиатора,- как правило, увеличивая его продолжительность. В-третьих, более крупные корковые нейроны получают множество возбуждающих и тормозных влияний от местных сетей с циркулирующим возбуждением, а также имеют множество различных типов рецепторов. Активация одного типа аминергического рецептора может привести к сильному или слабому эффекту в зависимости от исходного возбужденного состояния нейрона.
Наши знания о физиологии и фармакодинамике аминергических нейронов далеко не полные, однако их значение в широком разнообразии поведенческих функций не вызывает сомнений.
Часть поперечного среза через верхний отдел моста, показаны элементы ретикулярной формации.
Крупные, средние и мелкие нейроны сосредотачиваются в ядрах ретикулярной формации: субталамическом, красном, черной субстанции, мостовом, ретикулярных ядрах продолговатого мозга и др. Более сотни ядер, располагающихся в срединной, медиальной и латеральной группе. В срединных и медиальных ядрах находятся крупные нервные клетки, а в латеральных - средние и мелкие нейроны. Аксоны крупных нейронов часто образуют бифуркации, разделяясь на два отростка. Причем один отросток имеет восходящее направление вплоть до клеток коры, другой - нисходящее - к нейронам мозгового ствола, мозжечка, спинного мозга. Благодаря такому делению возникаютретикуло-петальные связи с вышележащими нейронами иретикуло-фугальные – с ниже расположенными нервными клетками. Мелкие и средние клетки являются, в основном, ассоциативными нейронами.
На них замыкаются чувствительные волокна от афферентных клеток коры большого мозга, гипоталамуса, мозжечка, спинного мозга (спино-ретикулярный тракт). В свою очередь отростки ретикулярных нейронов вступают во множественные контакты, очевидно, со всеми нейронами головного и спинного мозга.
Клетки ретикулярной формации входят в состав всех жизненно важных нервных центров - дыхательного, сердечно-сосудистого, пищеварительного и многих других, связанных с необходимыми физиологическими отправлениями: созреванием и выходом половых клеток, мочеиспусканием, дефекацией, теплорегуляцией, стереотипными движениями. Ретикулярные нейроны имеют связи с ядрами пирамидных и экстрапирамидных, висцеральных проводящих путей.
Ретикулярные центры с нисходящим направлением импульсов для органов дыхательной, сердечно-сосудистой и пищеварительной системы работают через симпатический отдел. Восходящая часть ретикулярной формации активирует корковые нейроны, и ее окончания диффузно разбросаны по всей коре.
В функциональном отношении ретикулярную формацию рассматривают:
как своеобразный “генератор энергии”, способный активизировать и поддерживать в рабочем состоянии все остальные нейроны (П.К. Анохин);
как регулятор функционального состояния нейронов (угнетение, активация, выключение);
как сложный рефлекторный центр, принимающий участие в контроле мышечного тонуса и стереотипных движений;
как центральную энцефалическую систему, отвечающую за работу всего мозга;
как биоэнергетическую систему всего организма;
как центр, определяющий и регулирующий волю человека.
Благодаря особенностям строения своих нейронов (бифуркация отростков, восходящие и нисходящие аксоны, многочисленные синапсы и др.) ретикулярная формация обеспечивает:
сохранность автоматизма дыхания и сердечных сокращений, температурного постоянства, глотания, мочеиспускания, дефекации, благодаря интегративной работе по объединению нейронов в различных отделах мозга и обеспечению вегетативной регуляции за счёт координации эфферентных и афферентных сигналов в соответствующих центрах мозгового ствола
процессы восприятия и формирование эмоций, воли, памяти, внимания и обучения, благодаря регулирующим потокам сенсорных импульсов в кору и подкорковые структуры играет роль своеобразного вентильного механизма, определяющего уровень сознания;
активность и тонус мышц всех видов;
активность и тонус клеток коры: различные степени бодрствования и сна, усталости и подъема сил и эмоций.
контроль рефлекторной деятельности путём стимуляции или торможения мотонейронов передних рогов серого вещества спинного мозга и двигательных ядер черепных нервов ствола мозга.
В медиальной части продолговатого мозга расположен ингибиторный центр, стимуляция которого понижает тонус скелетных мышц, угнетает рефлексы и препятствует прохождению к мотонейронам любых импульсов со стороны коры большого мозга и ядер экстрапирамидной системы. Напротив, ретикулярная формация моста и среднего мозга активирует нервные клетки периферических двигательных центров.
Участие в эмоциональном восприятии чувствительных сигналов путём увеличения или уменьшения поступления афферентных импульсов к лимбической системе. Регулируя медиаторный обмен нейронов, или модулируя активность их рецепторов с помощью определённых лекарственных препаратов или немедикаментозных средств, можно, в случае необходимости, продлить состояние бодрствования и активизировать деятельность коры больших полушарий или наоборот - добиться сна.