En los albores del desarrollo de la vida en la Tierra, todas las formas celulares estaban representadas por bacterias. Aspiraron materia orgánica disuelta en el océano primario a través de la superficie del cuerpo.
Con el tiempo, algunas bacterias se han adaptado para producir materia orgánica a partir de materia inorgánica. Para hacer esto, utilizaron la energía de la luz solar. Surgió el primer sistema ecológico en el que estos organismos eran productores. Como resultado, apareció oxígeno en la atmósfera terrestre, liberado por estos organismos. Con su ayuda, puede obtener mucho mas energia y utilizar la energía adicional para complicar la estructura del cuerpo: dividir el cuerpo en partes.
Uno de los logros importantes de la vida es la separación del núcleo y el citoplasma. El núcleo contiene información hereditaria. Una membrana especial alrededor del núcleo permitió protegerlo contra daños accidentales. Según sea necesario, el citoplasma recibe órdenes del núcleo que dirigen la actividad vital y el desarrollo de la célula.
Los organismos en los que el núcleo está separado del citoplasma han formado un super reino de lo nuclear (entre ellos se incluyen plantas, hongos, animales).
Por lo tanto, la célula, la base de la organización de plantas y animales, surgió y se desarrolló en el curso de la evolución biológica.
Incluso a simple vista, y mejor aún con una lupa, se puede ver que la pulpa de una sandía madura se compone de granos muy pequeños o granos. Estas son células, los “ladrillos” más pequeños que forman los cuerpos de todos los organismos vivos, incluidos los vegetales.
La vida de una planta se lleva a cabo mediante la actividad combinada de sus células, que forman un todo único. Con la multicelularidad de las partes de la planta, existe una diferenciación fisiológica de sus funciones, la especialización de varias células en función de su ubicación en el cuerpo de la planta.
Una célula vegetal se diferencia de una célula animal en que tiene un caparazón denso que cubre el contenido interno por todos lados. La célula no es plana (como suele representarse), lo más probable es que parezca una vesícula muy pequeña llena de contenido mucoso.
Consideremos una célula como unidad estructural y funcional de un organismo. En el exterior, la celda está cubierta con una pared celular densa, en la que hay áreas más delgadas: poros. Debajo hay una película muy delgada, una membrana que cubre el contenido de la célula, el citoplasma. Hay cavidades en el citoplasma: vacuolas llenas de savia celular. En el centro de la célula o cerca de la pared celular hay un cuerpo denso: un núcleo con un nucleolo. El núcleo está separado del citoplasma por una envoltura nuclear. Los cuerpos pequeños, plástidos, se distribuyen por todo el citoplasma.
Estructura de la célula vegetal
| Organoide | Dibujo | Descripción | Función | Peculiaridades |
Pared celular o membrana plasmática | Incoloro, transparente y muy duradero | Permite que las sustancias entren y salgan de la célula. | La membrana celular es semipermeable. |
|
Citoplasma | Sustancia viscosa espesa | Todas las demás partes de la celda se encuentran en él. | Está en constante movimiento |
|
El núcleo (una parte importante de la célula) | Redondeado u ovalado | Asegura la transferencia de propiedades hereditarias a las células hijas durante la división. | Parte central de la celda |
|
Forma esférica o irregular | Participa en la síntesis de proteínas. | |||
 | Un reservorio separado del citoplasma por una membrana. Contiene jugo celular | Se acumulan nutrientes de reserva y productos de desecho que son innecesarios para la célula. | A medida que la célula crece, pequeñas vacuolas se fusionan en una gran vacuola (central). |
|
Plastidios | Cloroplastos | Aprovecha la energía de la luz del sol y crea orgánicos a partir de inorgánicos. | La forma de los discos delimitados del citoplasma por una doble membrana |
|
Cromoplastos | Formado como resultado de la acumulación de carotenoides. | Amarillo, naranja o marrón |
||
 | Leucoplastos | Plástidos incoloros | ||
Caparazón nuclear | Consta de dos membranas (exterior e interior) con poros | Separa el núcleo del citoplasma | Permite el intercambio entre el núcleo y el citoplasma. |
La parte viva de la célula es un sistema estructurado, ordenado y limitado por membranas de biopolímeros y estructuras de membranas internas que participan en un conjunto de procesos metabólicos y energéticos que mantienen y reproducen todo el sistema como un todo.
Una característica importante es que no hay membranas abiertas con extremos libres en la célula. Las membranas celulares siempre limitan las cavidades o áreas, cerrándolas por todos lados.

Un diagrama generalizado moderno de una célula vegetal.
Plasmalema(membrana celular externa) - película ultramicroscópica de 7,5 nm de espesor, que consta de proteínas, fosfolípidos y agua. Es una película muy elástica que se humedece bien con agua y recupera rápidamente su integridad después de un daño. Tiene una estructura universal, es decir, típica de todas las membranas biológicas. En las células vegetales, fuera de la membrana celular, hay una pared celular fuerte que proporciona soporte externo y mantiene la forma de la célula. Consiste en fibra (celulosa), un polisacárido insoluble en agua.
Plasmodesmata Las células vegetales son túbulos submicroscópicos que penetran en las membranas y están revestidos por una membrana plasmática, que pasa así de una célula a otra sin interrupción. Con su ayuda, se produce la circulación intercelular de soluciones que contienen nutrientes orgánicos. También se utilizan para transferir biopotenciales y otra información.
Poro llamados agujeros en la membrana secundaria, donde las células están separadas solo por la membrana primaria y la placa mediana. Las áreas de la membrana primaria y la lámina mediana que separan los poros adyacentes de las células adyacentes se denominan membrana de poro o película de cierre de poro. La película de cierre de los poros es penetrada por los túbulos plasmodesmeables, pero a través del orificio generalmente no se forma en los poros. Los poros facilitan el transporte de agua y solutos de una célula a otra. En las paredes de las células vecinas, por regla general, una contra la otra, se forman poros.
Membrana celular tiene una cubierta bien definida, relativamente gruesa, de naturaleza polisacárido. La membrana de la célula vegetal es producto de la actividad del citoplasma. El aparato de Golgi y el retículo endoplásmico participan activamente en su formación.

Estructura de la membrana celular
La base del citoplasma es su matriz, o hialoplasma, un complejo sistema coloidal incoloro y ópticamente transparente capaz de realizar transiciones reversibles de sol a gel. El papel más importante del hialoplasma es unir todas las estructuras celulares en un solo sistema y asegurar la interacción entre ellas en los procesos del metabolismo celular.
Hialoplasma(o matriz del citoplasma) constituye el entorno interno de la célula. Consiste en agua y varios biopolímeros (proteínas, ácidos nucleicos, polisacáridos, lípidos), de los cuales la mayor parte son proteínas de diversa especificidad química y funcional. El hialoplasma también contiene aminoácidos, monosacáridos, nucleótidos y otras sustancias de bajo peso molecular.
Los biopolímeros forman un medio coloidal con agua, que, según las condiciones, puede ser denso (en forma de gel) o más líquido (en forma de sol), tanto en todo el citoplasma como en sus zonas individuales. En el hialoplasma, varios orgánulos e inclusiones se localizan e interactúan entre sí y con el entorno del hialoplasma. Además, su ubicación suele ser específica para ciertos tipos de células. A través de la membrana bilipídica, el hialoplasma interactúa con el entorno extracelular. En consecuencia, el hialoplasma es un entorno dinámico y juega un papel importante en el funcionamiento de los orgánulos individuales y la actividad vital de las células en general.
Los orgánulos (orgánulos) son componentes estructurales del citoplasma. Tienen cierta forma y tamaño, son estructuras citoplasmáticas obligatorias de la célula. En su ausencia o daño, la célula generalmente pierde su capacidad de continuar existiendo. Muchos de los orgánulos son capaces de dividirse y reproducirse. Sus dimensiones son tan pequeñas que solo se pueden ver a través de un microscopio electrónico.
El núcleo es el orgánulo más visible y generalmente el más grande de la célula. Fue explorado en detalle por primera vez por Robert Brown en 1831. El núcleo proporciona las funciones metabólicas y genéticas más importantes de la célula. Tiene una forma bastante variable: puede ser esférica, ovalada, lobulada, lenticular.
El núcleo juega un papel importante en la vida de la célula. La célula de la que se extrajo el núcleo ya no segrega membrana, deja de crecer y sintetizar sustancias. Los productos de descomposición y destrucción aumentan en él, como resultado de lo cual muere rápidamente. No se produce la formación de un nuevo núcleo a partir del citoplasma. Los nuevos núcleos se forman solo por división o aplastamiento del antiguo.
El contenido interno del núcleo es la carolinfa (jugo nuclear), que llena el espacio entre las estructuras del núcleo. Contiene uno o más nucléolos, así como una cantidad significativa de moléculas de ADN conectadas a proteínas específicas: histonas.

Estructura del núcleo
El nucleolo, como el citoplasma, contiene principalmente ARN y proteínas específicas. Su función más importante es la de formar ribosomas, que realizan la síntesis de proteínas en la célula.
El aparato de Golgi es un organoide que se distribuye universalmente en todo tipo de células eucariotas. Es un sistema de varios niveles de sacos de membrana planos, que se engrosan a lo largo de la periferia y forman procesos vesiculares. Con mayor frecuencia se encuentra cerca del núcleo.

Aparato de Golgi
El aparato de Golgi incluye necesariamente un sistema de pequeñas vesículas (vesículas), que se desprenden de cisternas engrosadas (discos) y se ubican a lo largo de la periferia de esta estructura. Estas vesículas desempeñan el papel de un sistema de transporte intracelular de gránulos de sectores específicos y pueden servir como fuente de lisosomas celulares.
Las funciones del aparato de Golgi también consisten en la acumulación, separación y excreción fuera de la célula con la ayuda de burbujas de los productos de síntesis intracelular, productos de descomposición y sustancias tóxicas. Productos de la actividad sintética de la célula, así como diversas sustancias que ingresan a la célula desde medio ambiente a través de los canales del retículo endoplásmico, se transportan al aparato de Golgi, se acumulan en este organoide y luego, en forma de gotas o granos, ingresan al citoplasma y son utilizados por la propia célula o extraídos. En las células vegetales, el aparato de Golgi contiene enzimas para la síntesis de polisacáridos y el propio material polisacárido, que se utiliza para construir la pared celular. Se cree que participa en la formación de vacuolas. El aparato de Golgi lleva el nombre del científico italiano Camillo Golgi, quien lo descubrió por primera vez en 1897.
Los lisosomas son pequeñas vesículas unidas por una membrana cuya función principal es realizar la digestión intracelular. El uso del aparato lisosómico ocurre durante la germinación de la semilla de la planta (hidrólisis de los nutrientes de reserva).
Estructura del lisosoma
Los microtúbulos son estructuras supramoleculares de membrana que consisten en glóbulos de proteínas dispuestos en hileras en espiral o rectilíneas. Los microtúbulos realizan principalmente una función mecánica (motora), proporcionando movilidad y contractilidad de los orgánulos celulares. Ubicados en el citoplasma, le dan a la célula una determinada forma y aseguran la estabilidad de la disposición espacial de los orgánulos. Los microtúbulos facilitan el movimiento de los orgánulos hacia lugares determinados por las necesidades fisiológicas de la célula. Un número importante de estas estructuras se encuentran en el plasmalema, cerca de la membrana celular, donde participan en la formación y orientación de las microfibrillas de celulosa de las membranas celulares vegetales.

Estructura de microtúbulos
La vacuola es la más importante componente células vegetales. Es una especie de cavidad (depósito) en la masa del citoplasma, llena con una solución acuosa de sales minerales, aminoácidos, ácidos orgánicos, pigmentos, carbohidratos y separada del citoplasma por una membrana vacuolar: tonoplasto.
El citoplasma llena toda la cavidad interna solo en las células vegetales más jóvenes. Con el crecimiento de la célula, la disposición espacial de la masa inicialmente continua de citoplasma cambia significativamente: aparecen pequeñas vacuolas llenas de jugo celular y toda la masa se vuelve esponjosa. Con un mayor crecimiento celular, las vacuolas individuales se fusionan, empujando las capas de citoplasma hacia la periferia, como resultado de lo cual generalmente hay una gran vacuola en la célula formada, y el citoplasma con todos los orgánulos se encuentra cerca de la membrana.
Los compuestos orgánicos y minerales solubles en agua de las vacuolas determinan las propiedades osmóticas correspondientes de las células vivas. Esta solución de cierta concentración es una especie de bomba osmótica para la penetración regulada en la célula y la liberación de agua, iones y moléculas de metabolitos de la misma.
En combinación con una capa de citoplasma y sus membranas, que se caracterizan por sus propiedades de semipermeabilidad, la vacuola forma un sistema osmótico eficaz. Indicadores de células vegetales vivas como el potencial osmótico, la fuerza de succión y la presión de turgencia se determinan osmóticamente.

Estructura de la vacuola
Los plástidos son los orgánulos citoplasmáticos más grandes (después del núcleo), inherentes solo a las células de los organismos vegetales. No se encuentran solo en los hongos. Los plastos juegan un papel importante en el metabolismo. Están separados del citoplasma por una membrana de doble membrana, y algunos de sus tipos tienen un sistema de membranas internas bien desarrollado y ordenado. Todos los plástidos son del mismo origen.
Cloroplastos- los plástidos de organismos fotoautótrofos más extendidos y funcionalmente más importantes, que llevan a cabo procesos fotosintéticos que finalmente conducen a la formación de materia orgánica y la liberación de oxígeno libre. Los cloroplastos de plantas superiores tienen una estructura interna compleja.

Estructura de cloroplasto
Tamaños de cloroplasto en diferentes plantas no son iguales, pero su diámetro promedio es de 4-6 micrones. Los cloroplastos pueden moverse bajo la influencia del movimiento del citoplasma. Además, bajo la influencia de la iluminación, hay un movimiento activo de cloroplastos de tipo ameba hacia la fuente de luz.
La clorofila es la sustancia principal de los cloroplastos. Gracias a la clorofila, las plantas verdes pueden utilizar energía luminosa.
Leucoplastos(plástidos incoloros) son cuerpos citoplasmáticos claramente marcados. Su tamaño es algo menor que el tamaño de los cloroplastos. Cada vez más monótonas y su forma, más cercana a la esférica.
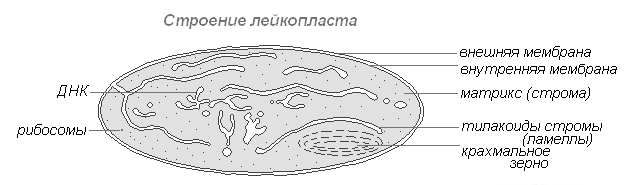
Estructura de leucoplastos
Se encuentra en las células de la epidermis, tubérculos, rizomas. Cuando se iluminan, se convierten muy rápidamente en cloroplastos con un cambio correspondiente en la estructura interna. Los leucoplastos contienen enzimas con la ayuda de las cuales el almidón se sintetiza a partir del exceso de glucosa formado en el proceso de fotosíntesis, la mayor parte del cual se deposita en tejidos u órganos de almacenamiento (tubérculos, rizomas, semillas) en forma de granos de almidón. En algunas plantas, las grasas se depositan en leucoplastos. La función de reserva de los leucoplastos se manifiesta ocasionalmente en la formación de proteínas de almacenamiento en forma de cristales o inclusiones amorfas.
Cromoplastos en la mayoría de los casos, son derivados de cloroplastos, ocasionalmente, leucoplastos.

Estructura de cromoplasto
La maduración de escaramujos, pimientos y tomates se acompaña de la transformación de cloro o leucoplastos de células pulpares en carotenoidoplastos. Estos últimos contienen principalmente pigmentos plástidos amarillos: carotenoides, que, cuando están maduros, se sintetizan intensamente en ellos, formando gotitas de lípidos de colores, glóbulos sólidos o cristales. En este caso, la clorofila se destruye.
Las mitocondrias son orgánulos característicos de la mayoría de las células vegetales. Tienen una forma variable de palos, granos, hilos. Descubierto en 1894 por R. Altman utilizando un microscopio óptico, y la estructura interna se estudió posteriormente con un microscopio electrónico.

Estructura de la mitocondria
Las mitocondrias tienen una estructura de dos membranas. La membrana externa es lisa, la interna forma excrecencias de varias formas: túbulos en las células vegetales. El espacio dentro de la mitocondria está lleno de un contenido semilíquido (matriz), que incluye enzimas, proteínas, lípidos, sales de calcio y magnesio, vitaminas, así como ARN, ADN y ribosomas. El complejo enzimático de las mitocondrias acelera el mecanismo complejo e interconectado de reacciones bioquímicas que dan como resultado la formación de ATP. En estos orgánulos, las células reciben energía: la transformación de la energía de los enlaces químicos de los nutrientes en enlaces ATP de alta energía en el proceso de respiración celular. Es en las mitocondrias donde se produce la descomposición enzimática de los carbohidratos, ácidos grasos, aminoácidos con la liberación de energía y su posterior conversión en energía ATP. La energía acumulada se gasta en procesos de crecimiento, en nuevas síntesis, etc. Las mitocondrias se multiplican por fisión y viven unos 10 días, después de lo cual se destruyen.
El retículo endoplásmico es una red de canales, túbulos, vesículas, cisternas ubicadas dentro del citoplasma. Descubierto en 1945 por el científico inglés K. Porter, es un sistema de membranas con estructura ultramicroscópica.
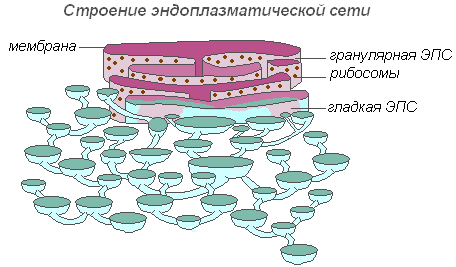
La estructura del retículo endoplásmico.
Toda la red está integrada en un solo todo con el externo membrana celular caparazón nuclear. Distinguir entre EPS liso y rugoso, con ribosomas. Los sistemas enzimáticos involucrados en el metabolismo de las grasas y los carbohidratos se encuentran en las membranas del EPS liso. Este tipo de membrana predomina en las células de las semillas ricas en sustancias de almacenamiento (proteínas, carbohidratos, aceites), los ribosomas se adhieren a la membrana del EPS granular y, durante la síntesis de una molécula de proteína, la cadena polipeptídica con ribosomas se sumerge en el canal del EPS. Las funciones del retículo endoplásmico son muy diversas: transporte de sustancias tanto dentro de la célula como entre células vecinas; división de la célula en secciones separadas, en las que se producen simultáneamente varios procesos fisiológicos y reacciones químicas.
Los ribosomas son orgánulos celulares sin membrana. Cada ribosoma consta de dos partículas que no son idénticas en tamaño y se pueden dividir en dos fragmentos, que continúan conservando la capacidad de sintetizar proteínas después de combinarse en un ribosoma completo.

Estructura del ribosoma
Los ribosomas se sintetizan en el núcleo, luego lo abandonan y pasan al citoplasma, donde se adhieren a la superficie externa de las membranas del retículo endoplásmico o se ubican libremente. Dependiendo del tipo de proteína sintetizada, los ribosomas pueden funcionar solos o combinarse en complejos: polirribosomas.
Formado con la participación del plasmalema. Es una formación extracelular multicapa que protege la superficie celular, siendo, de hecho, su esqueleto exterior. Consta de dos estructuras: una estructura (hecha de hilos de fibrillas de celulosa) y una matriz similar a un gel. La matriz contiene polisacáridos: hemicelulosas y sustancias pectínicas. Las hemicelulosas son cadenas poliméricas ramificadas que constan de varias hexosas (glucosa, manosa, galactosa) y pueden existir pentosas (xilosa, arabinosa) y ácidos urónicos (glucurónico y galacturónico). Estos componentes de las hemicelulosas se combinan entre sí en diferentes proporciones cuantitativas y forman varias combinaciones. Pero las cadenas de moléculas de hemicelulosa nunca cristalizan y las fibrillas no se fijan (no detectan) en ellas. Contiene una gran cantidad de agua.
Pectinas Grupo heterogéneo, que incluye polímeros ramificados altamente hidrogenados que portan cargas negativas debido a la presencia de muchos residuos de ácido galacturónico.
Gracias a estos componentes, la matriz es una masa plástica blanda, que es la base (marco).
Los polímeros de celulosa individuales se empaquetan en microfibrillas utilizando enlaces de hidrógeno. Aporta rigidez y fuerza. Entre ellos hay ligamentos que transforman la pared celular en un monolito. Estas microfibrillas de celulosa unidas también están rodeadas de pectinas. Pueden interactuar con calcio, silicio. Lo que le da rigidez. Debido a su estructura similar a un gel, la matriz proporciona una transmisión difusa de agua y moléculas pequeñas.
Las paredes celulares se dividen en 3 tipos: primaria, secundaria, terciaria.
La célula primaria está compuesta por un 90% de carbohidratos. Durante la división celular, aparece un haz de microtúbulos en el plano ecuatorial de las células, ubicado entre los cromosomas divergentes (phragmoplast). Entre los microtúbulos, hay muchas vesículas pequeñas del sistema vacuolar, CG; en la parte central, los fragmoplastos comienzan a fusionarse entre sí. Formando un disco aplanado o placa mediana. Consiste en un polisacárido - kalese. Más elástica en comparación con la celulosa (no está contenida dentro de las burbujas KG, se forma en la membrana plasmática). Las vesículas KG contienen los componentes necesarios para construir las membranas de dos células hijas. El proceso de fusión de pequeñas vacuolas ocurre desde el centro de la célula hacia la periferia y continúa hasta que las vesículas de la membrana se fusionan y se fusionan con la superficie de la superficie lateral de la célula.
La pared celular primaria en crecimiento ya consta de 3 capas (la lámina media, que consta de una matriz amorfa, y 2 periféricas, que contienen hemicelulosa y fibrillas). La membrana primaria está formada por la liberación de hemicelulosa y fibrillas de celulosa por dos nuevas estructuras celulares. Se producirá un aumento del grosor de la pared intercelular debido a la actividad de las células hijas, que, cada una por su parte, segregarán las sustancias de la membrana celular, engrosándose al superponerse cada vez más capas. Las fibrillas de celulosa se sintetizan y polimerizan en la membrana plasmática, lo que conduce a la formación de una pared celular secundaria (le da a la célula su forma final). Tan pronto como se completa el engrosamiento de la pared, se modifica con lignina, se sintetiza un polímero hidrófobo mediante polimerización oxidativa de hasta tres residuos de alcoholes aromáticos y sirve como los componentes principales de la madera (20-35% en madera - lignina). Debido a su naturaleza hidrófoba, la lignina crea impermeabilización en las paredes y sirve como material de refuerzo adicional. La pared celular se lignifica o se tapona (debido a la cutina y la suberina). En las células de la epidermis, la cera se libera en la superficie de las membranas celulares.
Los hongos tienen quitina (N-acetilglucosamina).
La pared celular (el término "pared celular" se usa a menudo como sinónimo del término "pared celular" en la literatura educativa y científica). Protoplasto.
La pared celular de la planta resiste la alta presión osmótica de la gran vacuola central y evita la ruptura celular. Además, el conjunto de fuertes paredes celulares desempeña el papel de una especie de esqueleto externo que mantiene la forma de la planta y le confiere resistencia mecánica. La pared celular, que posee una gran fuerza, es al mismo tiempo capaz de crecer y, sobre todo, de crecer por estiramiento. Estos dos, en cierta medida, requisitos opuestos se satisfacen debido a las peculiaridades de su estructura y composición química.
La pared celular suele ser transparente y transmite bien la luz solar. El agua y las sustancias de bajo peso molecular penetran fácilmente a través de él, pero para las sustancias de alto peso molecular es total o parcialmente impermeable. En los organismos multicelulares, las paredes de las células vecinas se mantienen unidas por sustancias de pectina que forman una placa mediana.
Cuando los tejidos vegetales se tratan especialmente con determinadas sustancias (álcalis fuertes, ácido nítrico), las paredes de las células vecinas se separan como resultado de la destrucción de la placa media. Este proceso se llama maceración. La maceración natural se produce en frutos demasiado maduros de pera, melón, melocotón, etc.
Como resultado de la presión de turgencia, las paredes de las células vecinas en las esquinas se pueden redondear y se forman espacios intercelulares entre ellas.
La pared celular es un producto de desecho de su protoplasto. Por lo tanto, la pared puede crecer solo cuando está en contacto con el protoplasto. Sin embargo, cuando el protoplasto muere, la pared se conserva y la célula muerta puede seguir desempeñando las funciones de transporte de agua o desempeñando el papel de soporte mecánico.
La base de la pared celular está formada por carbohidratos de alto polímero: moléculas de celulosa (fibra) ensambladas en haces complejos - fibrillas, que forman un marco inmerso en la base (matriz), que consta de hemicelulosas, pectinas y glicoproteínas (Fig.21) . Las moléculas de celulosa consisten en una gran cantidad de monómeros dispuestos linealmente: residuos de glucosa. La celulosa es muy estable, no se disuelve en ácidos diluidos e incluso en álcalis concentrados. El esqueleto de celulosa elástica imparte resistencia mecánica a la membrana celular. Inicialmente, el número de microfibrillas formadas por moléculas de celulosa en la pared celular es relativamente pequeño, pero con la edad aumenta y la célula pierde su capacidad de estirarse.
Las hemicelulosas se diferencian de la celulosa en la composición de los monómeros y su disposición ramificada en moléculas. Como uno de los componentes de la matriz plástica, las hemicelulosas dan resistencia adicional a la pared celular, pero casi no interfieren con su crecimiento. Las hemicelulosas también pueden ser sustancias de reserva, ya que se hidrolizan fácilmente. Además de las hemicelulosas, las sustancias de pectina o pectinas y los polisacáridos formados por monómeros, los ácidos urónicos, se incluyen en la matriz, así como en la placa central. Estas sustancias se mantienen unidas, pegan las cáscaras de las células vecinas. Las moléculas de hemicelulosas, pectina y glicoproteínas conectan las microfibrillas de celulosa.
Además de los polisacáridos, los componentes que no son carbohidratos se encuentran a menudo en la matriz de muchas paredes celulares. El más común de ellos es la lignina, una sustancia polimérica de naturaleza polifenólica. Su contenido en las paredes de algunos tipos de células puede llegar al 30%.
La membrana celular es un componente típico de una célula vegetal, producto de la vida del protoplasto.
Funciones:
1. Las membranas celulares fuertes y rígidas sirven como soporte mecánico para los órganos de las plantas.
2. La membrana limita el estiramiento del protoplasto por la vacuola, y el tamaño y la forma de la célula madura dejan de cambiar.
3. En los tejidos externos, las membranas celulares protegen las células más profundas para que no se sequen.
4. A lo largo de las paredes celulares adyacentes entre sí, varias sustancias y agua pueden moverse de una célula a otra (camino a través del apoplasto).
5. Influyen en la absorción, transpiración y secreción.
Las paredes de las células suelen ser incoloras y permiten que la luz del sol pase fácilmente. Las paredes de las células vecinas se sujetan con pectina. lámina mediana... La placa mediana es una capa única común a dos celdas adyacentes. Es una placa celular algo modificada que ha surgido en el proceso de citocinesis. La lámina media está menos regada; pueden estar presentes moléculas de lignina. Como resultado de la presión intracelular, las esquinas de las paredes celulares se pueden redondear y se forman espacios intercelulares entre las células adyacentes. Todas las paredes de las células vegetales, conectadas entre sí y adyacentes a los espacios intercelulares llenos de agua, aseguran la existencia de un ambiente regado continuo en el que las sustancias solubles en agua se mueven libremente.
Estructura y composición química.
Pared celular primaria.
Originalmente, surge fuera del plasmalema. celular primario pared.
Composición: celulosa, hemicelulosa, pectina y agua.
Las paredes celulares primarias de las células vecinas están conectadas por una placa mediana de protopectina. En la pared celular, las moléculas de celulosa lineales, muy largas (varias micras), que consisten en glucosa, se recolectan en haces (micelas, que, a su vez, se combinan en microfibrillas), las fibrillas más delgadas (1.5 ... 4 nm) de una forma indefinida. de longitud, y luego en macrofibrillas ... La celulosa forma un armazón multidimensional, que se sumerge en una matriz amorfa muy regada de carbohidratos no celulósicos: pectinas, hemicelulosas, etc. Es la celulosa la que aporta la fuerza de la pared celular. Las microfibrillas son elásticas y tienen una resistencia a la tracción similar al acero. Los polisacáridos de la matriz determinan propiedades de la pared tales como alta permeabilidad al agua, pequeñas moléculas e iones disueltos y un fuerte hinchamiento. Gracias a la matriz, el agua y las sustancias pueden moverse de una celda a otra a lo largo de las paredes adyacentes entre sí (el camino a través del apoplasto a través del "espacio libre"). Algunas hemicelulosas pueden depositarse en las paredes de las células de las semillas como sustancias de almacenamiento.
Crecimiento de la pared.
Cuando las células se dividen, solo se crea de nuevo la placa celular. Sobre él, ambas células hijas colocan sus propias paredes, que consisten principalmente en hemicelulosa. En este caso, la formación de la pared también se produce en la superficie interna de las paredes restantes que pertenecen a la célula madre. La lámina celular se transforma en una lámina mediana, suele ser delgada y casi indistinguible. Después de la división, la célula entra en la fase de estiramiento debido a la absorción de agua por la célula y al crecimiento de la vacuola central. La presión de turgencia estira la pared en la que se incrustan las micelas de celulosa y las sustancias de la matriz. Este método de crecimiento se llama intususcepción, implementación. Las membranas de las células en división y crecimiento se denominan primarias. Contienen hasta un 90% de agua, en materia seca predominan los polisacáridos de matriz: en pectinas dicotiledóneas y hemicelulosa en proporciones iguales, en monocotiledóneas, principalmente hemicelulosa; el contenido de celulosa no supera el 30%. El espesor de la pared primaria no supera los 0,1 ... 0,5 micrones.
Para cuando termina el crecimiento de la célula, el crecimiento de la pared celular puede continuar, pero ya en espesor. Este proceso se llama espesamiento secundario. En este caso, se deposita una pared celular secundaria sobre la superficie interior de la pared celular primaria. El crecimiento de la pared celular secundaria se produce como resultado aposiciones, superposición de nuevas micelas de celulosa en la superficie interna de la pared celular. Por tanto, las capas más jóvenes de la pared celular están más cerca del plasmallema.
Para algunos tipos de células (muchas fibras, traqueidas, segmentos vasculares), la formación de una pared secundaria es la función principal del protoplasto; una vez completado el engrosamiento secundario, muere. Sin embargo, esto no es requerido. La pared secundaria funciona principalmente de forma mecánica, funciones de apoyo... Contiene mucha menos agua y está dominada por microfibrillas de celulosa (40 ... 50% de materia seca). En las paredes secundarias de fibras de lino y cabellos de algodón, el contenido de celulosa puede alcanzar el 95%.
Mecanismo de construcción de la pared celular. La pared celular se forma como resultado de la actividad del protoplasto. De acuerdo con esto, las sustancias ingresan a la pared desde el interior, desde el lado del protoplasto. Materiales de construcción- moléculas de celulosa pectina, lignina y otras sustancias - se acumulan y se sintetizan parcialmente en los tanques del aparato de Golgi. Envasados en viales de Golgi, se transportan a la membrana plasmática. Habiéndolo roto, la burbuja estalla y su contenido queda fuera del plasmalema. La membrana de burbujas restaura la integridad de la membrana plasmática. Debido a la actividad enzimática del plasmalema, se ensamblan las fibrillas de celulosa y la estructura de la pared celular. Las fibrillas formadas por el plasmalema se aplican desde el interior sin entrelazar. En su orientación, los microtúbulos ubicados debajo del plasmalema en paralelo a las fibrillas en formación juegan un papel importante.
2. Poros. Modificaciones de la pared celular.
Poros. Cuando se forma la pared celular primaria, se distinguen áreas más delgadas en ella, donde las fibrillas de celulosa se encuentran más sueltas. Los túbulos de la cadena endoplásmica pasan a través de las paredes celulares aquí, conectando las células adyacentes. Estos sitios se llaman campos de poros primarios , y los túbulos del retículo endoplásmico que los atraviesan - plasmodesmos .
El crecimiento en espesor ocurre en la pared celular de manera desigual, pequeñas áreas de la pared celular primaria permanecen sin engrosar en las ubicaciones de los campos de poros primarios (canales de poros). Los canales de los poros de dos células vecinas suelen estar ubicados uno frente al otro y están separados por una película de poro de cierre: dos paredes celulares primarias con una sustancia intercelular entre ellas. La película retiene orificios submicroscópicos a través de los cuales pasan los plasmodesmos. Por lo tanto, el tiempo es dos canales de poros y una película de cierre entre ellos.
Los plasmodesmos penetran en las películas de los poros que se cierran. Cada célula contiene de varios cientos a decenas de miles de plasmodesmas. Los plasmodesmos se encuentran solo en las células vegetales, donde hay paredes celulares sólidas. Los plasmodesmos se forman a partir de túbulos ER que permanecen en la lámina celular entre dos células hijas. Cuando se recrea el RE de ambas células, se conectan a través de los plasmodesmos.
Plasmodesmata pasa a través del canal de plasmodesma en la película de cierre del poro. El plasmalema que recubre el conducto y el hialoplasma entre éste y el plasmodesma son continuos con los plasmalemas y el hialoplasma de las células adyacentes. Por tanto, los protoplastos de las células vecinas están interconectados por los canales de plasmodesmos y plasmodesmos. A través de ellos se produce un transporte intercelular de iones y moléculas, además de hormonas. Los protoplastos de las células de una planta unidas por plasmodesmos forman un todo único: un simplastos. El transporte de sustancias a través de los plasmodesmos se denomina simplástico, en contraste con el transporte apoplásico a lo largo de las paredes celulares y los espacios intercelulares.
En el proceso de la vida celular, la pared celular de celulosa puede sufrir modificaciones.
Pared celular (pared celular)- un rasgo característico de una célula vegetal que la distingue de una célula animal. La pared celular le da a la célula una forma específica. Las células vegetales cultivadas en medios nutritivos especiales, de los que se elimina la pared enzimáticamente, siempre adoptan una forma esférica. La pared celular da fuerza celular y protege el protoplasto, equilibra la presión de turgencia y así evita la ruptura del plasmalema. La colección de paredes celulares forma el esqueleto interno que sostiene el cuerpo de la planta y le da fuerza mecánica.
La pared celular es incolora y transparente, transmite fácilmente la luz solar. Por lo general, las paredes están saturadas de agua. El transporte de agua y compuestos de bajo peso molecular disueltos en ella (transporte a lo largo del apoplasto) se realiza a lo largo del sistema de paredes celulares.
La pared celular se compone principalmente de polisacáridos, que se pueden subdividir en materia esquelética y sustancias de la matriz.
Sustancia esquelética la pared celular vegetal es celulosa (fibra), que es beta-1,4-D-glucano. Es la materia orgánica más abundante de la biosfera. Las moléculas de celulosa son cadenas no ramificadas muy largas, están dispuestas paralelas entre sí en grupos de varias decenas y se mantienen unidas por numerosos enlaces de hidrógeno. Como resultado, microfibrillas, que crean el marco estructural de la pared y determinan su resistencia. Las microfibrillas de celulosa son visibles solo en un microscopio electrónico, su diámetro es de 10-30 nm, la longitud alcanza varias micras.
La celulosa es insoluble y no se hincha en agua. Es químicamente inerte, insoluble en disolventes orgánicos, álcalis concentrados y ácidos diluidos. Las microfibrillas de celulosa son elásticas y muy resistentes al desgarro (similar al acero). Estas propiedades determinan el uso generalizado de la celulosa y sus productos. La producción mundial de fibra de algodón, que se compone casi en su totalidad de celulosa, es de 1,5 10 7 toneladas por año. De la celulosa se obtienen polvos sin humo, acetato de seda y viscosa, celofán y papel. Se lleva a cabo una reacción cualitativa a la celulosa con un reactivo. yodo de zinc de cloro, la pared celular de celulosa se tiñe de azul violeta.
En los hongos, la sustancia esquelética de la pared celular es quitina- un polisacárido construido a partir de residuos de glucosamina. La quitina es incluso más duradera que la celulosa.
Las microfibrillas se sumergen en amorfo matriz, generalmente un gel plástico saturado de agua. La matriz es una mezcla compleja de polisacáridos, cuyas moléculas están compuestas por residuos de varios azúcares diferentes y son más cortas que las de celulosa y cadenas ramificadas. Los polisacáridos de la matriz determinan propiedades de la pared celular como un fuerte hinchamiento, alta permeabilidad al agua y compuestos de bajo peso molecular disueltos en ella, y propiedades de intercambio catiónico. Los polisacáridos de la matriz se dividen en dos grupos: sustancias de pectina y hemicelulosa.
Sustancias de pectina hincharse fuertemente o disolverse en agua. Son fácilmente destruidos por álcalis y ácidos. Los representantes más simples de las sustancias de pectina son solubles en agua. ácidos pécticos- productos de polimerización del ácido alfa-D-galacturónico (hasta 100 unidades) unidos por enlaces 1,4 en cadenas lineales (alfa-1,4-D-galacturonano). Ácidos pécticos (pectinas)- estos son compuestos poliméricos de mayor peso molecular (100-200 unidades) de ácido alfa-D-galacturónico, en los que los grupos carboxilo están parcialmente metilados. Pectatos y pectinados- Sales de calcio y magnesio de los ácidos péctico y péctico. Los ácidos pécticos, pectatos y pectinatos son solubles en agua en presencia de azúcares y ácidos orgánicos para formar geles densos.
Las paredes celulares de las plantas contienen principalmente protopectinas- polímeros de alto peso molecular de ácido poligalacturónico metoxilado con arabinos y galactanos; en plantas dicotiledóneas, las cadenas de galacturonano contienen una pequeña cantidad de ramnosa. Las protopectinas son insolubles en agua.
Hemicelulosa son cadenas ramificadas construidas a partir de residuos de azúcares neutros, glucosa, galactosa, manosa, xilosa son más comunes; grado de polimerización 50-300. Las hemicelulosas son químicamente más estables que las sustancias pectínicas, son más difíciles de hidrolizar y se hinchan menos en agua. Las hemicelulosas se pueden depositar en las paredes de las células de las semillas como sustancias de reserva (palmera datilera, caqui). Las sustancias pectínicas y las hemicelulosas están unidas por transiciones mutuas. Además de los polisacáridos, en la matriz de las paredes celulares está presente una proteína estructural especial. Está asociado con los residuos del azúcar arabinosa y, por tanto, es una glicoproteína.
Los polisacáridos de matriz hacen más que simplemente llenar los espacios entre las microfibrillas celulósicas. Sus cadenas están dispuestas de manera ordenada y forman numerosos enlaces tanto entre sí como con microfibrillas, lo que aumenta significativamente la fuerza de la pared celular.
Las paredes celulares de las plantas a menudo se modifican químicamente. Lignificación, o lignificación ocurre si la matriz se deposita lignina- Compuesto polimérico de naturaleza fenólica, insoluble en agua. La pared celular lignificada pierde su elasticidad, su dureza y resistencia a la compresión aumentan bruscamente y su permeabilidad al agua disminuye. Los reactivos para la lignina son: 1) floroglucinol y clorhídrico concentrado o ácido sulfurico(las paredes lignificadas adquieren un color rojo cereza) y 2) sulfato anilina, bajo la influencia de la cual las paredes lignificadas se vuelven de color amarillo limón. La lignificación es característica de las paredes celulares del tejido conductor del xilema (madera) y del tejido mecánico del esclerénquima.
Suberización, o suberinización ocurre como resultado de la deposición de polímeros hidrofóbicos en el lado interno de la pared celular - suberina y cera... Suberin es una mezcla de ésteres de ácidos grasos poliméricos. Los monómeros de cera son alcoholes grasos y ésteres de cera. La cera se elimina fácilmente con disolventes orgánicos y se funde rápidamente y forma cristales. Suberin es un compuesto amorfo que no se funde ni se disuelve en disolventes orgánicos. Suberin y cera, formando capas paralelas alternas, recubren toda la cavidad celular desde el interior en forma de película. La película de suberina es prácticamente impermeable al agua y a los gases, por lo que, después de su formación, la célula suele morir. El taponamiento es característico de las paredes celulares del tejido integumentario del corcho. El reactivo en la pared celular tapada con corcho es SudánIII, color rojo anaranjado.
Cutinización expuesto a las paredes externas de las células del tejido tegumentario de la epidermis. Kutin y cera depositado en capas alternas en la superficie exterior de la pared celular en forma de película - cutículas... Kutin es un compuesto polimérico similar a la grasa, similar en naturaleza química y propiedades a la suberina. La cutícula protege a la planta de la evaporación excesiva del agua de la superficie de la planta. Puedes pintarlo con un reactivo. SudánIII en color rojo anaranjado.
Mineralización La pared celular se produce debido a la deposición en la matriz. un número grande minerales, más a menudo sílice (óxido de silicio), menos a menudo oxalato y carbonato de calcio. Los minerales imparten dureza y fragilidad a la pared. La deposición de sílice es característica de las células epidérmicas de colas de caballo, juncos y cereales. La rigidez de tallos y hojas adquirida como resultado de la silicificación sirve como agente protector contra los caracoles y también reduce significativamente la comestibilidad y el valor nutricional de las plantas.
Algunas células especializadas tienen paliza pared celular. En este caso, en lugar de una pared secundaria de celulosa, se depositan polisacáridos ácidos amorfos altamente hidratados en forma moco y chicle que son similares en naturaleza química a las sustancias de pectina. El moco se disuelve bien en agua con la formación de soluciones mucosas. Encías pegajosas, estiradas en hilos. Cuando se secan, tienen una consistencia córnea. Cuando se deposita moco, el protoplasto se empuja gradualmente hacia el centro de la célula, su volumen y el volumen de la vacuola disminuyen gradualmente. Eventualmente, la cavidad celular puede llenarse completamente con moco y la célula muere. En algunos casos, el moco puede atravesar la pared celular primaria hacia la superficie. El aparato de Golgi participa principalmente en la síntesis y secreción de moco.
El moco secretado por las células vegetales tiene varias funciones. Entonces, el moco del casquete de la raíz sirve como lubricante para facilitar el crecimiento de la punta de la raíz en el suelo. Las glándulas mucosas de las plantas insectívoras (rocío del sol) secretan moco atrapante, al que se adhieren los insectos. El moco secretado por las células externas de la cubierta de la semilla (lino, membrillo, plátanos) fija la semilla a la superficie del suelo y evita que la plántula se seque. El moco se tiñe con un reactivo. azul de metileno en azul.
La liberación de encías suele ocurrir cuando las plantas se lesionan. Por ejemplo, la fuga de goma de las áreas heridas de troncos y ramas se observa a menudo en cerezas y ciruelas. El pegamento de cereza es una goma solidificada. La encía realiza una función protectora, cubriendo la herida desde la superficie. Las encías se forman principalmente en plantas leñosas de las familias de leguminosas (acacia, tragacanto astrágalo) y subfamilias de ciruela rosácea (cereza, ciruela, albaricoque). Las encías y el moco se utilizan en medicina.
La pared celular es un producto de desecho del protoplasto. Los polisacáridos de la matriz, la glicoproteína de la pared, la lignina y el moco se forman en el aparato de Golgi. La síntesis de celulosa, la formación y orientación de microfibrillas son realizadas por el plasmalema. Un papel importante en la orientación de las microfibrillas pertenece a los microtúbulos, que se encuentran paralelos a las microfibrillas depositadas cerca del plasmalema. Si se destruyen los microtúbulos, solo se forman células isodiamétricas.
La formación de la pared celular comienza durante la división celular. En el plano de división, se forma una placa celular, una sola capa común a dos células hijas. Consiste en sustancias pectínicas de consistencia semilíquida; sin celulosa. En una célula adulta, la placa celular se conserva, pero sufre cambios, por lo que se llama mediana, o lámina intercelular (sustancia intercelular) (arroz. 2.16). La placa mediana suele ser muy delgada y casi indistinguible.
Inmediatamente después de la formación de la placa celular, los protoplastos de las células hijas comienzan a depositar su propia pared celular. Se deposita desde el interior tanto en la superficie de la placa celular como en la superficie de otras paredes celulares que anteriormente pertenecían a la célula madre. Después de la división, la célula entra en la fase de crecimiento por estiramiento, que se debe a la intensa absorción osmótica de agua por la célula asociada con la formación y crecimiento de la vacuola central. La presión de turgencia comienza a estirar la pared, pero no se rompe debido al hecho de que nuevas porciones de microfibrillas y sustancias de la matriz se depositan constantemente en ella. La deposición de nuevas porciones de material se produce de manera uniforme en toda la superficie del protoplasto, por lo que el grosor de la pared celular no disminuye.
Las paredes de las células en división y crecimiento se denominan primario... Contienen mucha (60-90%) de agua. En materia seca predominan los polisacáridos de matriz (60-70%), el contenido de celulosa no supera el 30% y no hay lignina. El espesor de la pared primaria es muy pequeño (0,1-0,5 micrones).
Para muchas células, la deposición de la pared celular se detiene simultáneamente con el cese del crecimiento celular. Estas células están rodeadas por una delgada pared primaria hasta el final de su vida ( arroz. 2.16).
Arroz. 2.16. Célula parenquimatosa con pared primaria.
En otras celdas, la deposición de la pared continúa incluso después de que la celda alcanza su tamaño final. En este caso, el grosor de la pared aumenta y el volumen ocupado por la cavidad de la celda disminuye. Este proceso se llama engrosamiento secundario paredes, y la pared en sí se llama secundario(arroz. 2.17).
La pared secundaria puede considerarse como una función de soporte adicional, principalmente mecánica. Es la pared secundaria la responsable de las propiedades de la madera, fibras textiles, papel. La pared secundaria contiene significativamente menos agua que la primaria; está dominado por microfibrillas de celulosa (40-50% del peso de materia seca), que se encuentran paralelas entre sí. De los polisacáridos de la matriz, las hemicelulosas (20-30%) son características, hay muy pocas sustancias pectínicas. Las paredes celulares secundarias suelen estar lignificadas. En las paredes secundarias no lignificadas (fibras de líber de lino, pelos de algodón), el contenido de celulosa puede alcanzar el 95%. El alto contenido y la orientación estrictamente ordenada de las microfibrillas determinan las altas propiedades mecánicas de las paredes secundarias. A menudo, las células con una pared celular secundaria lignificada mueren después de que se completa el engrosamiento secundario.
La lámina mediana pega las células adyacentes. Si se disuelve, las paredes celulares pierden su conexión entre sí y se desconectan. Este proceso se llama maceración. La maceración natural es bastante común, en la que las sustancias pectínicas de la placa central se convierten en un estado soluble con la ayuda de la enzima pectinasa y luego se lavan con agua (frutos demasiado maduros de pera, melón, melocotón, plátano). A menudo se observa una maceración parcial, en la que la placa media se disuelve no en toda la superficie, sino solo en las esquinas de las celdas. Debido a la presión de turgencia, las células adyacentes en estos lugares se redondean, como resultado de lo cual espacios intercelulares(arroz. 2.16). Los espacios intercelulares forman una única red ramificada, que está llena de vapor de agua y gases. Así, los espacios intercelulares mejoran el intercambio de gases de las células.
Un rasgo característico del muro secundario es su deposición desigual sobre el muro primario, como resultado de lo cual quedan áreas no engrosadas en el muro secundario - poros... Si la pared secundaria no es muy gruesa, los poros parecen depresiones poco profundas. En las celdas con una pared secundaria potente, los poros de la sección tienen la forma de canales radiales que se extienden desde la cavidad celular hasta la pared primaria. Por la forma del canal de poros, se distinguen poros de dos tipos: sencillo y sobre afilado(figura 2.17).

Arroz. 2.17. Tipos de poros: A - células con paredes secundarias y numerosos poros simples; B - un par de poros simples; B - un par de poros bordeados.
Tengo poros simples el diámetro del canal de poros a lo largo de toda la longitud es el mismo y tiene la forma de un cilindro estrecho. Los poros simples son característicos de las células parenquimatosas, estopa y fibras de madera.
Los poros en dos celdas adyacentes tienden a aparecer uno frente al otro. Estos poros comunes tienen la forma de un solo canal, separados por un tabique delgado de la placa media y la pared primaria. Tal conjunto de dos poros de paredes adyacentes de células vecinas se llama pares de poros y funciona como uno. La sección del muro que los separa se llama película de cierre poros, o membrana de poro... En las células vivas, la película de cierre de los poros está impregnada de numerosos plasmodesmos(arroz. 2.18).
Plasmodesmata son inherentes solo a las células vegetales. Son cordones de citoplasma que atraviesan la pared de las células adyacentes. La cantidad de plasmodesmas en una célula es muy grande: desde varios cientos hasta decenas de miles, generalmente los plasmodesmas se recolectan en grupos. El diámetro del canal de plasmodesma es de 30 a 60 nm. Sus paredes están revestidas con plasmalema, continuo con el plasmalema de las células adyacentes. En el centro del plasmodesmo hay un cilindro de membrana: barra central plasmodesmos continua con las membranas de los elementos del retículo endoplásmico de ambas células. Entre el bastón central y el plasmalema, hay un hialoplasma en el canal, que se continúa con el hialoplasma de las células adyacentes.

Arroz. 2.18. Plasmodesmata bajo un microscopio electrónico (esquema): 1 - en un corte longitudinal; 2 - en una sección transversal; Pl- plasmalema; California- la barra central del plasmodesma; ER- un elemento del retículo endoplásmico.
Por lo tanto, los protoplastos de las células no están completamente aislados entre sí, sino que se comunican a través de los canales de los plasmodesmos. A través de ellos se produce un transporte intercelular de iones y pequeñas moléculas, así como estímulos hormonales. A través de los plasmodesmos, los protoplastos de las células de un organismo vegetal forman un todo único, llamado symplast, y el transporte de sustancias a través de los plasmodesmos se llama simplástico diferente a apoplásico transporte a lo largo de las paredes celulares y los espacios intercelulares.
Tengo poros bordeados(arroz. 2.17) el canal se estrecha bruscamente en el proceso de deposición de la pared celular, por lo tanto, la abertura interna del poro, que entra en la cavidad celular, es mucho más estrecha que la externa, apoyándose contra la pared primaria. Los poros con bordes son característicos de las células que mueren temprano de los elementos de la madera que conducen el agua. En ellos, el canal de poros se expande a modo de embudo hacia la película de cierre, y la pared secundaria cuelga en forma de cresta sobre la parte expandida del canal, formando una cámara de poros. El nombre del poro bordeado proviene del hecho de que, visto desde la superficie, la abertura interior parece un círculo pequeño o una rendija estrecha, mientras que la abertura exterior parece bordear la interior en forma de un círculo de mayor diámetro o hendidura más ancha.
Los poros facilitan el transporte de agua y solutos de una célula a otra sin comprometer la resistencia de la pared celular.